")
Ein systemdynamischer Designansatz
Aufgrund der Bedeutung, im Kontext moderner Aquaponik, haben wir diese Studie auf Deutsch übersetzt. Für Fehler in der Übersetzung übernehmen wir keine Haftung. Hier das Original: Navigating towards Decoupled Aquaponic Systems
Simon Goddek 1,2,*, Carlos Alberto Espinal 3 , Boris Delaide 4, Mohamed Haissam Jijakli 4, Zala Schmautz 5, Sven Wuertz 6 und Karel J. Keesman 1
1 Biobased Chemistry and Technology, Wageningen University, PO Box 17, Wageningen 6700 AA, Niederlande;
2 Aquaponik Manufaktur GmbH, Geldener Str. 139, Issum 47661, Deutschland
3 LandIng Aquaculture, Evenheuvel 4, 5688 LZ Oirschot, Niederlande;
4 Integriertes und städtisches Pflanzenpathologielabor, Université de Liége, Avenue Maréchal Juin 13, Gembloux 5030, Belgien;
5 Institut für Natürliche Ressourcen, Zürcher Hochschule für Angewandte Wissenschaften (ZHAW), Grüental, Wädenswil 8820, Schweiz;
6 IGB, Ökophysiologie und Aquakultur, Müggelseedamm 310, 12587 Berlin, Deutschland;
* Korrespondenz:
Fachredakteurin: Giuliana Parisi; Eingegangen: 24. Februar 2016; Angenommen: 12. Juli 2016; Veröffentlicht: 21. Juli 2016
Zusammenfassung : Das klassische Funktionsprinzip der Aquaponik besteht darin, nährstoffreiches Aquakulturwasser
einer hydroponischen Pflanzenzuchtanlage zuzuführen, die das Wasser anschließend reinigt und in die
Aquakulturbecken zurückführt. Ein bekannter Nachteil ist, dass für die Produktion von Nutzpflanzen und Fischen unter denselben Umweltbedingungen Kompromisse hinsichtlich der optimalen Wachstumsbedingungen eingegangen werden müssen . Ziel dieser Studie war die Entwicklung eines theoretischen Konzepts für ein entkoppeltes Aquaponiksystem (DAPS) und die Vorhersage von Wasser-, Nährstoff- (N und P), Fisch-, Schlamm- und Pflanzenkonzentrationen. Hierzu wurde ein dynamisches Aquaponiksystemmodell entwickelt, das auf Daten aus der Literatur zu Aquakultur, Hydroponik und Schlammbehandlung basiert. Die Modellergebnisse zeigten die Abhängigkeit der Aquakulturwasserqualität von der Evapotranspirationsrate der Hydroponik. Dieses Ergebnis lässt sich dadurch erklären, dass DAPS auf unidirektionalen Flüssen basiert. Diese unidirektionalen Nährstoffflüsse führen zur Anreicherung remineralisierter Nährstoffe in der Hydrokultur und gewährleisten so optimale Wachstumsbedingungen für die Pflanzen. Die Studie empfiehlt außerdem, die Anbaufläche anhand der Phosphorverfügbarkeit in der Hydrokultur zu dimensionieren, da Phosphor eine endliche Ressource ist und als einer der wichtigsten limitierenden Faktoren für das Pflanzenwachstum identifiziert wurde . Schlüsselwörter: Aquaponik; Aquakulturtechnik; Klärschlammbehandlung; Landwirtschaft; anaerobe Vergärung; Phosphatrecycling
1. Einleitung.
Aquaponik kann als integriertes, quasi geschlossenes, multitrophisches Lebensmittelproduktionssystem definiert werden, bestehend aus einem Kreislauf-Aquakultursystem (RAS) und einer Hydrokulturanlage. In einem Aquaponiksystem wird das nährstoffreiche Wasser aus der Fischzucht in die Hydrokulturanlage geleitet und versorgt die Pflanzen mit Nährstoffen. Der Reiz der Aquaponik liegt in ihrer Fähigkeit, Wassertiere (z. B. Fische, Krebse usw.) und Pflanzen (z. B. Gemüse, Kräuter, Heilpflanzen, Früchte usw.) umweltfreundlich zu produzieren und dabei eine hohe Wassernutzung [1] und Nährstoffrückführung [2,3] zu gewährleisten. Trotz unterschiedlicher Anforderungen an die Wasserqualität in RAS und Hydrokulturanlagen beruht die traditionelle Aquaponikpraxis darauf, die Bedürfnisse von Pflanzen und Fischen in einem einzigen Wasserkreislauf zu berücksichtigen. Dies reduziert die Effizienz von Aquaponiksystemen im Vergleich zur
Summe der Einzelproduktion von Nutzpflanzen, Fischen bzw. Pflanzen [4]. Obwohl viele Aquaponiksysteme als Kreislaufsysteme konzipiert und betrieben werden, entwickeln kommerzielle Anbauer und Forscher diese Systeme kontinuierlich weiter, um eine unabhängige Steuerung jeder einzelnen Systemkomponente
(z. B. Kreislaufanlage, Hydrokultur und Nährstoffrückgewinnung durch Schlammremineralisierung) zu ermöglichen. Systeme, in denen Fische, Pflanzen und gegebenenfalls die Remineralisierung als separate Funktionseinheiten mit jeweils eigenem Wasserkreislauf integriert sind und unabhängig voneinander gesteuert werden können, werden als entkoppelte Aquaponiksysteme (DAPS) bezeichnet. Ziel der Systementwicklung ist dennoch ein hoher Grad an Selbstversorgung des
Gesamtsystems. Die Komponenten werden daher so ausgelegt und dimensioniert, dass der Aufwand für die Anpassung der Kreislaufbedingungen minimiert wird. Kürzlich stellten Kloas et al. [5] ein DAPS vor, das aus einer Kreislaufanlage und einer Hydrokultureinheit besteht, die als zwei separate Wasserkreisläufe angeordnet sind. Der durch Evapotranspiration der Pflanzen verursachte Wasserverlust wird bei Bedarf über ein Rückschlagventil der Kreislaufanlage ausgeglichen,
die anschließend mit Leitungswasser aufgefüllt wird. Dadurch wurde eine verbesserte Steuerung der Nährstoffflüsse sowie eine Optimierung der artspezifischen Wasserbedingungen in beiden Einheiten erreicht. Der entscheidende Faktor dieses Ansatzes ist der Wasserverbrauch (d. h. hauptsächlich die Evapotranspirationsrate), da er den Wasseraustausch und die Wasserqualität im Kreislaufsystem sowie die Nährstoffversorgung der Pflanzen bestimmt, sofern keine zusätzliche Düngung erfolgt. Daher ist das Verständnis der Auswirkungen von Wasser- und Nährstoffflüssen innerhalb solcher Systeme unerlässlich für die Festlegung ihres konzeptionellen Rahmens. Wachstumsvorteile von DAPS wurden in Laborexperimenten beobachtet [6,7]. Jijakli et al. [6] stellten ein um 39 % gesteigertes Pflanzenwachstum im Vergleich zu einer reinen hydroponischen Kontrollnährlösung fest, nachdem die hydroponische Komponente mit zusätzlichem Dünger angereichert wurde. Auch Goddek [7] zeigte, dass anaerobe Gärreste das Pflanzenwachstum förderten. Unseres Wissens wurde das DAPS-Konzept bisher nicht auf Systeme mit mehr als zwei Einheiten angewendet. In dieser Studie erweiterten wir das Konzept durch die Integration einer dritten Funktionseinheit zur Remineralisierung von RAS-Schlamm. Ähnlich einem Nahrungsnetz könnten verschiedene Funktioneinheiten, die unterschiedliche trophische Ebenen repräsentieren – wie autotrophe Produzenten (Nutzpflanzen), heterotrophe Konsumenten (Fische, Krebstiere, Weichtiere) und Destruenten (Remineralisierungseinheit) – in einzelnen, aber miteinander verbundenen Wasserkreisläufen angeordnet werden. Dies ermöglicht eine bessere Kontrolle der Nährstoffflüsse, steigert die Produktionseffizienz und reduziert die Emissionen weiter. Am wichtigsten ist, dass dieses Konzept eine präzisere Steuerung der Nährstoffflüsse zwischen verschiedenen Einheiten ermöglicht und die Stabilität gegenüber Ungleichgewichten oder Störungen erhöht.
Die Nährstoffdynamik in integrierten Aquakultur-Hydroponik-Systemen mit einem Kreislauf wurde erstmals von Seawright et al. [8] untersucht. Aufgrund nicht dokumentierter Nährstoffzugaben blieben Nährstoffflüsse und -ansammlungen jedoch unklar. Graber und Junge [2] sowie Licamele [9] führten eine Stickstoffbilanzstudie für Aquaponik-Systeme mit einem Kreislauf durch und beobachteten, dass das Wachstum von Salat in Hydrokultur und Aquaponik ähnlich war, obwohl der Salat in der Hydrokultur einer Nährlösung mit höherer Konzentration ausgesetzt war. Neto und Ostrensky [10] führten eine weitere Massenbilanzstudie durch und schätzten die Nährstoffbelastung und die Abfallproduktion in Tilapia-Käfighaltungssystemen. Bezüglich des Nährstoffflusses von der Kreislaufanlage zu den Pflanzen in Ein-Kreislauf -Systemen haben mehrere Autoren dieses Thema untersucht und niedrige Gehalte an Phosphor, Kalium, Magnesium, Mangan, Eisen und Schwefel festgestellt [1,2,9,11–13]. Folglich könnte das Pflanzenwachstum wahrscheinlich durch die Zufuhr ausreichender Nährstoffe verbessert werden. Dies kann entweder durch Supplementierung [4,14] oder durch Remineralisierung von Fischschlamm erreicht werden, wie in mehreren Studien mit Massenbilanzansätzen beobachtet wurde [15–17].
2. Ziele und Anwendungsbereich
Modelle werden typischerweise erstellt, um komplexe dynamische Systeme zu verstehen, vorherzusagen und zu steuern [18].
Ziel dieser Studie ist die Entwicklung eines Systemdynamikmodells für Stickstoff (N), Phosphor (P), Wasser, Fische, Pflanzen und Klärschlamm (Bioschlamm aus der Aquakultur) auf Basis eines entkoppelten Systemansatzes. Der Makronährstoff Phosphor wird in dieser Arbeit verwendet, da er eine erschöpfliche Ressource ist [19].
Obwohl er im Fischfutter größtenteils ausreichend vorhanden ist, können andere, besser verfügbare Nährstoffe der Hydrokulturkomponente zugeführt werden. Das Systemdynamikmodell dieser Studie kann zur Auslegung und weiteren Optimierung von Systemen eingesetzt werden. Hauptziel dieser Arbeit ist die Vorstellung eines theoretischen Auslegungsansatzes für DAPS (Digital Aquaponic Systems), indem die konzeptionellen Kriterien des Anwendungsbereichs berücksichtigt und Auslegungsschwächen auf Basis der Modellergebnisse behandelt werden. Das in dieser Arbeit entwickelte Modell bewertet die organischen Belastungsraten des Systems, um unter gegebenen Bedingungen optimale Bedingungen für die Hydrokultur- und die RAS-Komponente (Rezirkulations- und Kreislaufanlage) zu erreichen. Dies führt zu einer Perspektive, die die Bewertung des Bedarfs an weiteren Studien umfasst, die zur Entwicklung und Verbesserung eines pflanzenweiten Modells für entkoppelte Aquaponiksysteme durchgeführt werden müssen.
3. Methodik und Datenquellen
3.1. Dynamische Systemanalyse
Die dynamische Systemanalyse wurde eingesetzt, um die Stickstoff-, Phosphor-, Fisch- (d. h. Tilapia-), Pflanzen- (d. h. Salat-) und Wasserbilanz innerhalb des DAPS zu bewerten. Diese Methode ermöglichte es uns auch, den Einfluss einer Remineralisierungskomponente auf die Gesamtleistung des Systems zu beurteilen und den hydroponischen Teil in Abhängigkeit von verschiedenen Evapotranspirationsraten zu dimensionieren. Als Grundlage für physikalisch basierte dynamische Modellierung – im Gegensatz zur datengetriebenen Modellierung – sind jedoch fundierte Flussdiagramme und Kausaldiagramme (CLD) erforderlich [18]. Während Flussdiagramme einen Überblick über alle im Rahmen eines umfassenden Diagnoseprozesses als notwendig erachteten Schritte bieten, stellen CLDs ein grundlegendes Werkzeug zum Verständnis und zur Veranschaulichung komplexer Systeme dar. Beide dienten als Grundlage und bildeten die Basis für das Software-Computermodell.
Obwohl kausale Flussdiagramme (CLDs) ein gutes Werkzeug zur Identifizierung von Kausalzusammenhängen zwischen Aktivitäten oder Ereignissen und
ihren latenten Auswirkungen darstellen, müssen sie mit dem Faktor Zeit kombiniert werden, um kurz- bis langfristige Einflussfaktoren und Veränderungen, z. B. aufgrund von Akkumulationen, Temperaturschwankungen und Fischbiomasse, zu simulieren und aufzuzeigen. Die resultierenden Systemdynamikmodelle ermöglichen die Analyse des Zusammenspiels von Schlüsselfaktoren, um wichtige Hebelpunkte sowie optimale Bedingungen und Systemeinstellungen zu ermitteln. Hier wurde die spezifische Systemmodellierungssoftware AnyLogic verwendet [20]. Für die CLDs (siehe Abbildungen B1–B3)
wurden folgende Annahmen getroffen:
(1) RAS, Hydroponik und Schlammremineralisierung werden unabhängig voneinander dargestellt, um den Bedarf an unterschiedlichen Bedingungen in jedem Teilsystem abzubilden; und (2) in der RAS-Komponente ist ein nitrifizierender Biofilter im Modell enthalten. Als Grundlage für die Modelle
gelten folgende Annahmen für die Flussdiagramme: (1) Es besteht kein zusätzlicher Wasseraustausch über den durch Evapotranspiration bedingten Wasserersatz hinaus;
(2) Es wird vorausgesetzt, dass die RAS-Komponenten Biofilter ausreichender Größe enthalten, um den gesamten Ammoniakstickstoff (TAN) vollständig in Nitrat umzuwandeln; und (3) Nährstoffzugaben für die Hydrokultureinheiten werden nicht berücksichtigt. Die Simulationen des Systemdynamikmodells basierten auf einem entkoppelten System mit vier Fischbecken. Das Modellierungsverfahren war in vier Schritte unterteilt: (1) Bevor die Flüsse von Stickstoff (N), Phosphor (P), Wasser, Tilapia, Salat und Schlamm simuliert wurden, wurde ein Parametervariationsexperiment durchgeführt, um die Fischmenge zu ermitteln, die für eine maximale Besatzdichte von 50 kg/m³ in das System eingebracht werden muss.3. Bei höheren Pflanzendichten kann die Verwendung von reinem Sauerstoff erforderlich sein. (2) Basierend auf diesen Daten wurde die Anbaufläche für die maximale Stickstoff-Nitrat-Konzentration (N-NO₃) im Wasser für empfindliche und resistente Fischarten (z. B. Forelle und Tilapia) mittels eines Parametervariationsexperiments in Abhängigkeit von der Evapotranspirationsrate (ETc) der Pflanzen unter natürlichen und künstlichen Lichtbedingungen simuliert. Für Forellen wurde der Grenzwert auf 40 mg × L⁻¹ N-NO₃ [ 21–23], für Tilapia auf 200 mg × L⁻¹ N -NO₃ festgelegt .
(3) Phosphor (P) wurde als alternativer Auslegungsparameter für die Dimensionierung der Hydrokulturkomponente verwendet, da er häufig das Pflanzenwachstum begrenzt und die globalen Mineralressourcen endlich sind. Sollte die ermittelte Dimensionierungsempfehlung jedoch nicht den erforderlichen Wasseraustauschraten entsprechen, ist eine aktive Denitrifikation im Kreislaufsystem erforderlich. Für diese Studie wurde ein Optimierungsschritt durchgeführt, um die Menge an Salat zu bestimmen, die mit Nährstoffen aus dem Kreislaufsystem (RAS) und remineralisiertem Phosphor aus einer anaeroben Nährstoffremineralisierungsanlage (ANRC) produziert werden kann. Die ANRC bestand in dieser Studie aus einem UASB-Reaktor (Upflow Anaerobic Sludge Blanket). Die Remineralisierungsraten wurden anhand der Leistungsparameter des UASB-Reaktors ermittelt ;
(4) Ausgehend vom Phosphorparameter wurde der Durchfluss des UASB bestimmt.
3.2. FAO-Penman-Monteith-Gleichung
Die Evapotranspirationsrate hängt von der Nettostrahlung, der Temperatur, der Windgeschwindigkeit, der relativen Luftfeuchtigkeit und der Pflanzenart ab [24, 25]. Die Nettosolarstrahlung kann mithilfe der FAO-
Penman-Monteith-Gleichung bestimmt werden [25]. Diese Gleichung wurde ursprünglich für Freilandbedingungen entwickelt, kann aber für den Anbau von Pflanzen in Gewächshäusern angepasst werden [24, 26]. Neben einer rein natürlichen Abhängigkeit haben sowohl die Eigenschaften des Gewächshauses als auch die Klimatisierungstechnik einen großen Einfluss auf die Evapotranspiration [27, 28]. Fernández et al. [29] berichten von einer Reduzierung der Referenzverdunstung (ETo) um 21,4 % bei der Verwendung von Kunststoffgewächshäusern in Spanien. Gewächshäuser reduzieren zudem die Windgeschwindigkeit erheblich,
was sich ebenfalls negativ auf die Referenzverdunstung auswirkt [29, 30]. Oberflächenabdeckungen , wie sie beispielsweise bei Nährstoffflusstechniken (NFT) oder Tiefwasserkulturen (DWC) eingesetzt werden , reduzieren den Einzelkulturkoeffizienten (Kc) [25], der mit der Referenzverdunstung multipliziert wird, um
die Evapotranspiration der Kulturpflanze zu ermitteln. Licamele [9] berichtete, dass eine Pflanzendichte von 32 Salatpflanzen pro m² jeweils 1 Liter Wasser benötigte. Diese Ergebnisse stimmten mit dem geschätzten Einzelkulturkoeffizienten für abgedeckte Flächen überein (Abbildung 1). Daher wurde mit AnyLogic eine vergleichende Analyse durchgeführt, um die Bandbreite der zu erwartenden Evapotranspirationsraten unter konstanten Beleuchtungsbedingungen von 200 W/ m² für 16 h/Tag und bei ausschließlicher Nutzung von natürlichem Licht zu schätzen.
3.2. FAO-Penman-Monteith-Gleichung
Die Evapotranspirationsrate hängt von der Nettostrahlung, der Temperatur, der Windgeschwindigkeit, der relativen Luftfeuchtigkeit und der Pflanzenart ab [24, 25]. Die Nettostrahlung kann mithilfe der FAO-Penman-Monteith-Gleichung bestimmt werden [25]. Diese Gleichung wurde ursprünglich für Freilandbedingungen entwickelt, kann aber für den Anbau von Pflanzen in Gewächshäusern angepasst werden [24, 26]. Neben einer rein natürlichen Abhängigkeit haben sowohl die Eigenschaften des Gewächshauses als auch die Klimatisierungstechnik einen großen Einfluss auf die Evapotranspiration [27, 28]. Fernández et al. [29] berichten von einer Reduzierung der Referenzverdunstung (ETo) um 21,4 % bei Verwendung von Kunststoffgewächshäusern in Spanien. Gewächshäuser reduzieren zudem die Windgeschwindigkeit erheblich, was sich ebenfalls negativ auf die Referenzverdunstung auswirkt [29, 30]. Es wurde berichtet, dass Oberflächenabdeckungen, wie sie beispielsweise bei Nährstoffflusstechniken (NFT) oder Tiefwasserkulturen (DWC) eingesetzt werden, den Pflanzenkoeffizienten (Kc) [25] reduzieren, der mit der Referenzverdunstung multipliziert wird, um die Pflanzenverdunstung zu ermitteln. Licamele [9] berichtete, dass eine Pflanzendichte von 32 Salatpflanzen pro Quadratmeter jeweils 1 Liter Wasser benötigte. Diese Ergebnisse stimmten mit dem geschätzten Pflanzenkoeffizienten für abgedeckte Flächen überein (Abbildung 1). Daher wurde mit AnyLogic eine vergleichende Analyse durchgeführt, um die Bandbreite der zu erwartenden Verdunstungsraten unter konstanten Lichtbedingungen von 200 W/m² für 16 h/Tag und bei ausschließlicher Nutzung von natürlichem Licht zu schätzen.
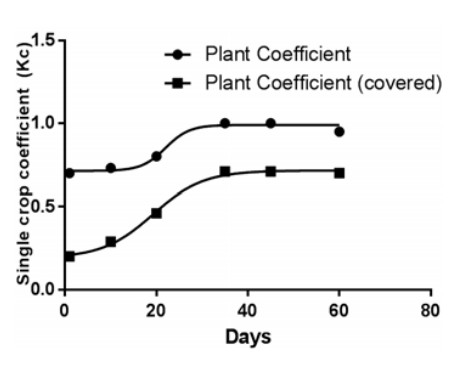
Abbildung 1. Anbaukoeffizient für Kopfsalat bei unbedecktem und oberflächenbedecktem Anbau unter der Annahme, dass
die Setzlinge nach 15 Tagen gepflanzt werden (im Diagramm als Tag 0 dargestellt).
Bei Kenntnis von ETo und Kc wurde die Evapotranspiration der Kulturpflanze in m³ pro m² x Tag - 1, üblicherweise ausgedrückt in mm x Tag - 1, wie folgt geschätzt:
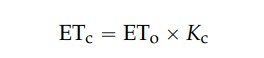
wobei ETc die Evapotranspiration der Kulturpflanze (mm Tag⁻¹) und Eto die Referenzevapotranspiration (mm Tag⁻¹ ) ist.
Kc ist der Pflanzenkoeffizient für Kopfsalat (dimensionslos). Die Referenzverdunstung für die natürliche Beleuchtung wurde anhand gemessener stündlicher Solarstrahlungsdaten für Köln-Bonn (Tabelle 1), Deutschland (50°47'N; 7°51'O) aus dem Jahr 2014 [31] geschätzt , die repräsentativ für die mitteleuropäische Region sind. Der Temperaturbereich im Gewächshaus wurde auf 20–24 °C festgelegt, die relative Luftfeuchtigkeit auf 60–80 %. Es wurde angenommen, dass die
Lichtdurchlässigkeit der Gewächshausverglasung die einfallende Strahlung um 10 % reduziert. Der Verschattungsfaktor durch die Konstruktion und die Umgebung reduzierte sie um weitere 15 %, wodurch angenommen wurde, dass die Nettostrahlung des natürlichen Lichts unter Gewächshausbedingungen um 25 % abnimmt. Abbildung 2 zeigt die geschätzte ETo-Differenz unter natürlicher und künstlicher Beleuchtung. Die Referenzverdunstung bei natürlicher Beleuchtung
kann durch die folgende Formel ausgedrückt werden, die sich auf die natürliche Beleuchtungskurve in Abbildung 2 bezieht,
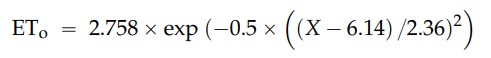 wobei X die Zeit in Monaten ist.
wobei X die Zeit in Monaten ist.
Kommentar hinzufügen